
神戸大学大学院農学研究科のハサン メエラジ (博士後期課程)と理化学研究所環境資源科学研究センター高橋聡史 (テクニカルスタッフ) らは、ハクサイにおいて、遺伝子発現調節に重要なヒストンの化学修飾※1 であるH3K4me3とH3K36me3の役割を明らかにしました。
本研究は、神戸大学大学院農学研究科の藤本龍准教授ら、理化学研究所環境資源科学研究センター関原明チームリーダーら、オーストラリア連邦科学産業研究機構 (CSIRO) らの研究グループにより行われました。
この研究成果は、2021年6月7日に、Frontiers in Plant Science にオンライン掲載されました。
ポイント
- ハクサイの遺伝子発現調節に重要なヒストン化学修飾であるヒストンH3の4番目のリジン残基のトリメチル化 (H3K4me3) とH3K36me3の役割を明らかにした。
- H3K36me3は遺伝子の発現の活性化や異なる組織間での恒常的な遺伝子発現に重要である。
- 転写活性型のヒストン化学修飾 (H3K4me3) と転写抑制型のヒストン化学修飾 (H3K27me3) の両方の修飾を持つBivalentクロマチン構造を構築している遺伝子を見出した。
研究の背景
DNA配列が同じ遺伝子を持つ個体間や同一個体内であっても、その遺伝子の発現レベルには違いが見られることが知られています。例えば、植物の同じ個体においても、葉、根、花といった異なる組織では、異なった遺伝子群が発現しています。この場合、各組織のDNA塩基配列は同じであることから、DNA配列以外の情報によって遺伝子発現が制御されていることになります。このようなDNA配列に依存しない遺伝子の発現制御はエピジェネティック※2 な制御とよばれています。エピジェネティックな修飾として、DNAメチル化やヒストンの化学修飾等が知られています。近年、DNA配列を決定する技術が発達しており、多くの生物種で全ゲノムDNA配列の解読が行われています。これに伴い、ゲノム全体のエピジェネティックな修飾パターン (ゲノムに対しエピゲノム※3 と呼ぶ) も、様々な生物種で明らかにされています。
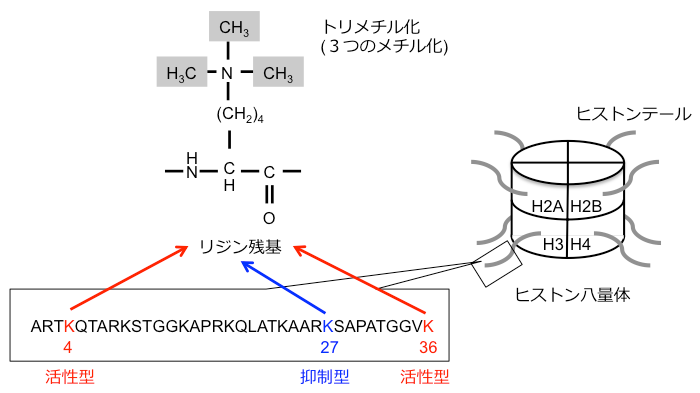
本研究の研究対象であるハクサイ (Brassica rapa L.) の全ゲノムDNA配列は2011年に決定されました。ゲノム全体のヒストンの化学修飾であるH3K27me3 (図1)の修飾状態については以前本研究チームが報告していますが、H3K4me3やH3K36me3 (図1)については報告がありませんでした。また、転写活性型のH3K4me3?H3K36me3と転写抑制型のH3K27me3の関係性についても報告がありませんでした。
研究の内容
ハクサイの2系統について、葉の組織を用いて、ゲノム全体でのH3K4me3とH3K36me3の修飾状態を明らかにしました。そして、H3K4me3とH3K36me3の有無と遺伝子発現量との関係について調べました。2系統でH3K4me3 (~17,000遺伝子)とH3K36me3 (~12,000遺伝子) 修飾を持つ遺伝子の発現量の平均値と、全遺伝子 (~40,000遺伝子) の発現量の平均値を比較しました。その結果、H3K4me3とH3K36me3の修飾を持つ遺伝子は全遺伝子の発現量の平均値に比べて、発現量が高くなり、特にH3K36me3を有する遺伝子でその傾向が見られました (図2)。このことは、H3K4me3とH3K36me3の修飾は遺伝子発現を活性化する役割があることを示します。さらに、H3K4me3とH3K36me3の修飾を持つ遺伝子は、様々な組織で発現レベルが変動しにくい (恒常的な遺伝子発現を示す) 傾向にあることも明らかになりました (図3)。
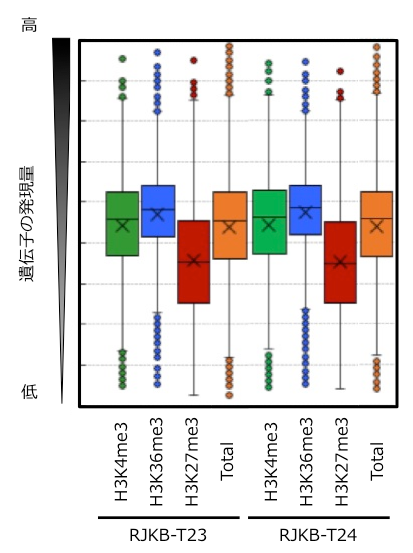
Totalは全遺伝子を示す。
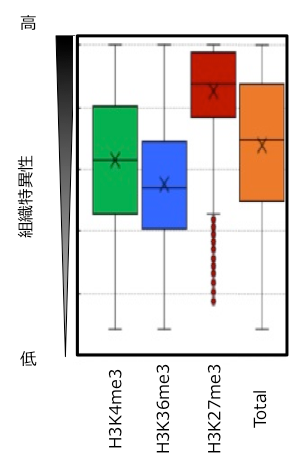
Totalは全遺伝子を示す。
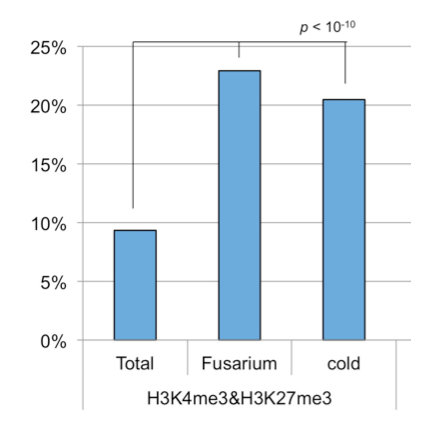
生物ストレス: 病原菌 (Fusarium) 感染後に発現が変動する遺伝子
環境ストレス: 4週間の低温 (cold) 処理後に発現が変動する遺伝子
Total: 全遺伝子に占めるH3K4me3とH3K27me3をもつ遺伝子の割合
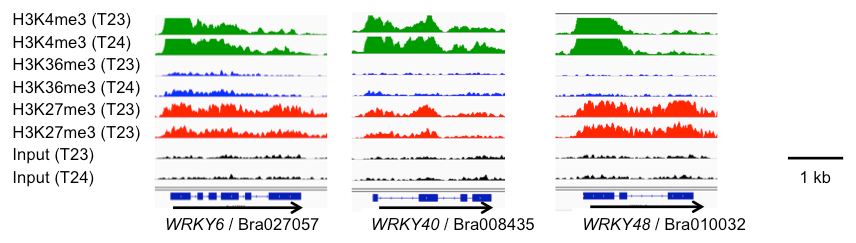
以前報告した転写抑制型のH3K27me3の結果を加えて、3つのヒストン化学修飾の関係性を調べた結果、H3K36me3とH3K27me3は互いに拮抗的な関係性にあることが明らかとなりました。また、H3K4me3とH3K27me3の両方の修飾を持つBivalentクロマチン構造を構築している可能性がある遺伝子群 (~3,700遺伝子) を見出しました (図4)。
Bivalentクロマチン構造を有する遺伝子群は、転写調節に関わる遺伝子に多く見られ、さらに、生物ストレスや環境ストレスによって発現が変動した遺伝子に多く含まれていたことから (図5)、Bivalentクロマチン構造によって、遺伝子発現の活性化と抑制化の両方に対応できる状態を構築している可能性が考えられました。実際に、幾つかの遺伝子について、Bivalentクロマチン構造を検出できる方法を用いて調べた結果、確かにBivalentクロマチン構造を構築していることが確認されました。
本研究から、ハクサイにおいて、転写活性型のヒストン化学修飾 (H3K4me3とH3K36me3) の遺伝子発現への役割、さらには、転写抑制型ヒストン化学修飾 (H3K27me3) との関係性について明らかにすることができました。
今後の展開
今回の研究から、ハクサイにおけるH3K4me3やH3K36me3修飾のゲノム全体での役割が明らかになりました。今後は個別の遺伝子におけるH3K4me3やH3K36me3の役割や、植物の形質への関わりを明らかにすることで、ハクサイの農業形質との関連性等について詳細に調べたいと考えています。また、転写活性型のH3K4me3と転写抑制型H3K27me3の両方の修飾を持つBivalentクロマチン構造を構築している遺伝子群を見出し、これらがストレス応答において重要な役割を担う可能性が示唆されました。今後は、これらの遺伝子についても個別に調べ、ハクサイのストレス応答との関わりについて調べたいと考えています。
用語解説
※1 ヒストンの化学修飾
ヒストンは核に存在するタンパク質で、4種類のコアヒストン (H2A、H2B、H3、H4) によってヒストン8量体を形成する。このヒストン8量体にDNAが巻き付いて、ヌクレオソームと呼ばれる染色体の最小単位を形成する。そして、複数のヌクレオソームがクロマチン構造を形成する。ヒストンの特定のリジン残基がアセチル化やメチル化され、クロマチン構造が変化し、遺伝子の発現が活性化されたり、抑制化されたりすることをヒストンの化学修飾という。
※2 エピジェネティック
DNAの塩基配列変化を伴わず、染色体の変化から生じる安定的に継承される遺伝子発現や形質のこと。
※3 エピゲノム
DNAの塩基配列変化を伴わない遺伝情報のこと。DNAメチル化やヒストンの化学修飾、非コードRNA、クロマチンの高次構造などを含んだ、遺伝子の働きを定める情報。
謝辞
本研究は、以下の支援を受けて行われました。
- 文部科学省科学研究費新学術領域研究(研究領域提案型)『生命科学系3分野支援活動』「ケ?ノム支援」(221S0002)
- 日本学術振興会 二国間共同研究事業
- 日本学術振興会 国際共同研究加速基金 (16KK0171)
- 日本学術振興会 科学研究費 若手A (15H05614)、基盤B (18H02173, 19H02947)
論文情報
タイトル
“Characterization of histone H3 lysine 4 and 36 tri-methylation in Brassica rapa L.”
DOI
10.3389/fpls.2021.659634
著者
Hasan Mehraj+, Satoshi Takahashi+, Naomi Miyaji, Ayasha Akter, Yutaka Suzuki, Motoaki Seki, Elizabeth S. Dennis, Ryo Fujimoto
+ These authors equally contributed to this work掲載誌
Frontiers in Plant Science










